Cyanobacteria




**
Colony usually microscopic, but sometimes macroscopic in a few species, subsphaerical in some plankton forms, but terrestrial species are usually irregular in shape. Colony with many irregularly arranged cells embedded in mucilage, with outer margin of which is usually rather indistinct. Cells sphaerical or subsphaerical, with binary division, but successive divisions perpendicular to each. The cells tend to be packed quite closely in forms associated with terrestrial or submerged plants, but are more spaced apart in some planktonic forms. No gas vacuoles.
Small genus comprising three planktic species known from eutrophic freshwaters. The genus was erected to group together colonial organisms characterised by vegetative cells that contain brownish-ring-shaped semi-terminal iron intrusions.
The main diacritical features of the genus include:
Cells small (1 – 2 × 0.5 – 1.5 μm), free-floating, solitary or in small colonial groups
Cells cylindrical, elongate with rounded ends
Cells with brownish or blackish ferric precipitates (on or around the cells)
Cell division in one plane in successive generations, transversely to the longitudinal axis of the cell, often where the precipitations are located.
**




**
**
**
Cells or small groups of cells surrounded by common mucilaginous envelopes, more commonly in microscopic, irregular or more or less spherical gelatinous colonies, usually with a small number of cells (2 – 16), rarely solitary, sometimes forming compound, many celled agglomerations.
Cells form within multi-cellular colonies sometimes in packet-like 2 – 8 celled groups. Mucilaginous envelopes colourless or slightly yellowish, often concentrically lamellate, delimited, and well defined at the margins.
Cells spherical, sub-spherical or widely oval, hemispherical after division; ± aerotopes. Cell division in at least 3 planes in successive generations usually perpendicular to one another; cells not reaching original size before division.

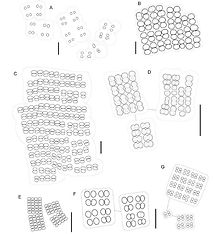


Colony planktonic, sphaerical or irregularly ellipsoid or oval mucilaginous mass, with cells forming an irregular layer some distance from the outside of the mucilage.Cells sphaerical or slightly enlongated in radial axis (but not heart shaped); at the ends of narrow, distinct, branched stalks originating near centre of colony. Cell division perpendicular to surface of colony, with successive divisions in two different planes. Cells are often spaced some distance from each other, and, even if quite close, never in direct contact.
**
Colony free-floating, sphaerical or subsphaerical, up to 100um diameter, with cells restricted to a zone near the surface, through a narrow layer of mucilage is always evident between the cells and the outside of the colony; cells in a layer mostly only one cell thick, though there are usually a few other cells just inside the layer. Cells sphaerical or subsphaerical, 2-4um diameter; gas vacuolate when in the plankton. It is important to view different planes within the colony, as a casual glance might suggest that these cells occur throughout.
Colonies free-floating, more or less globose or ellipsoidal, irregularly spherical, sometimes composed of sub-colonies.
Cells solitary, without aerotopes, elongated - reniform, arranged in a single layer at the surface of the colony, connected by structureless branched stands of hyaline mucilage.
Cell division in one plane perpendicular to the longitudinal axis in successive generations. Cells reach original size before next cell division. Reproduction by disintegration of colony.



**
Planktonic or benthic. Trichomes either solitary and generally straight or aggregated in fine tangled mats, isopolar, markedly constricted at the cross-walls, without branching, sometimes motile by slow gliding and trembling; firm shealths lacking but fine thin diffluent mucilaginous sheaths rare in some species. Cells isodiametric or mainly longer than wide, with or without aerotopes, when present these localised at the poles or in the centre of the cell; end cells sometimes slightly attenuated at the apex, without a calyptra, sometimes with characteristic apical protrusions. Reproduction by trichome fragmentation into many short few- celled motile hormogonia; necridia absent; all cells in the trichome capable of dividing and growing to +- original size before the next division.
Pseudanabaena catenata – Cosmopolitan species commonly found among decaying vegetation and debris in streams, ponds and reservoirs. African isolated are known to produce the neurotoxins cyanotoxin and anatoxin (Gorham et al., 1982).
Pseudanabaena galeata – Cosmopolitan species common in the plankton of lakes and reservoirs throughout Australia. Also found growing among benthic cyanoprokaryotes assembles in shallow littoral areas of rivers and wetlands; widely distributed in many biotopes throughout north-eastern Australia. Known to produce taste and odour compounds 2-methylisoborneol and Geosmin (Izaguirre & Taylor, 2004).
Pseudanabaena limnetica – Common in the plankton of lakes and reservoirs, also associated with the benthos. Found throughout north-eastern Australia, and considered to be cosmopolitan. Populations from Bjelke-Petersen Dam, south-east Queensland, regularly occur with acuminate apical cells and possibly represent a distinct species.
Pseudanabaena mucicola – Endogloeic within the mucilage of planktonic and benthic populations of colonial cyanoprokaryotes such as Microcystis, Aphanothece, Chroococcus, and Wollea. This species has a cosmopolitan distribution.
*Not known to be toxigenic in Australia. Some spp produce earthy, musty odours
Metabolite produced – MIB/geosmin
**
Characteristics: Cells of Microcystis are arranged in colonies that are
initially spherical, but become irregular or perforated over time. The cells may
be grouped tightly or sparsely within the fine, colourless colonial mucilage.
The mucilage is often not clearly seen in preserved material. Smaller
colonies are microscopic, while larger colonies may be viewed with the naked
eye. Each colony consists of thousands of very small individual cells that are
spherical to sub-spherical without individual mucilage sheaths. Although the
protoplast is a pale blue-green colour, the cells, when viewed through the light
microscope, often appear black as a result of gas vacuoles that are located
within the cells. The gas vacuoles allow the colony to drift through water
layers to find the optimal amount of sunlight. In some species the gas
vacuoles appear glistening or reddish because of the reflection of light.
Species are differentiated by, amongst others, cell-size, presence of gas
vacuoles, the nature of the sheath, and the shape of the colony.
Dimensions: Cells vary from 0.5-9 μm in diameter.
Ecology: Microcystis is usually part of the phytoplankton, but may also form
granular clumps on bottom substrates. Colonies are common in enriched
lakes, ponds and reservoirs or in slow-flowing eutrophic rivers. Microcystis,
like many other cyanobacteria, prefers high water temperatures and usually
form blooms during the summer periods under conditions of adequate
nutrient supply. When environmental conditions are favourable, large
numbers (blooms) can sometimes be seen floating on the surface of the
water, giving a blue-green tinge to the water. It is interesting that, where
blooms of some species (e.g. M. aeruginosa (Kützing) Kützing) occur, the
habitat is completely dominated by this species to the exclusion of almost all
other forms of cyanobacteria.
Notes: In contrast to M. aeruginosa colonies, which are highly irregular and
clathrate when mature, M. flos-aquae (Wittrock) Kirchner occurs in nearly
globular colonies. M. wesenbergii (Komárek) Komárek also has intensely
lobed colonies, but it can be distinguished from M. flos-aquae and
M. aeruginosa by a smooth, very firm and colourless mucilage with the outer
margin of the colony clearly delimited and extending 3-6 μm beyond the cell
aggregations. The presence of gas vacuoles (which appear black or dark
brown under the light microscope) easily distinguishes Microcystis from
Aphanocapsa Nägeli (which lacks gas vacuoles).
Problems: Microcystis is a common cause of algal blooms, sometimes
secreting chemicals that inhibit other algae. Because of the presence of gas
vacuoles that render them buoyant, they produce surface scums and cause
a great deal of disturbance in lakes and reservoirs. Dense growths may lead
directly or indirectly to the death of fish through suffocation (as a result of
oxygen depletion) or by poisoning. Microcystis can produce a polypeptide,
called microcystin (named after Microcystis), which is toxic to animals
ingesting contaminated water. It has also been implicated in human
illnesses, such as necrosis of the liver (from ingestion) and severe dermatitis
(from skin contact). Blooms of Microcystis can also impart taste and odour to
the water and interfere with recreational activities.
Microcystis blooms often form during warm, calm weather in lakes and ponds with relatively high nutrient concentrations (nitrogen or phosphorous) or low nitrogen to phosphorus ratio (N:P<15)
Recent work suggests that high total phosphorus (TP) or total Nitrogen (TN) concentrations are better predictors of bloom formation than N:P ratios.
Microcystis does not fix atmospheric nitrogen, but is very efficient in taking up ammonium and urea, and often blooms when nitrogen concentrations are limiting to other algae. Microcystis blooms may follow blooms of nitrogen fixing Aphanizomenon.
Microcystis can use low concentrations of the herbicide glyphosate as a phosphorous source. The gas vesicles in Microcystis cells provide a mechanism to move up and down in the water column, which increases access to nutrients and other growth factors.
Microcystis blooms usually contain other types of cyanobacteria, especially Aphanizonmenon, Dolichospermum, Gloeotrichia, and Woronichinia. Microcystis colonies may be surrounded by short filaments of Pseudanabaena and inactive Chlamydomonas cells.
Microcystis cells may produce anatoxins (nerve toxin), Microcystins (liver toxins), lipopolysaccharides (skin irritants), and BMAA ( beta-Methylamino-L-alanine; nerve toxin). These toxins are released into the ambient environment when the cell wall is disrupted (cell lysis).
The toxin microcystin was first isolated from Microcystis aeruginosa.
Microcystins are rapidly degraded by naturally occurring but specialised bacteria. If the specialised bacteria are not present, Microcystins can persist in the aquatic environment for months. Anatoxins are rapidly degraded by sunlight and at pH levels that are slightly above neutral. At low pH levels, and in the absence of light, anatoxins may persist in the aquatic environment for a few weeks.
BMAA can bioaccumulate in zooplankton and fish, so this nerve toxin can contribute to health risks long after the toxic bloom has died back. Higher water temperatures and light appear to be associated with increased toxin production.
Not all Microcystis blooms result in the release of toxins.
*Potentially toxic – Microcystins
Sulphurous, earthy
Metabolite produced – Beta cyclo-citral, iso-propyl mercaptan, disulphides (aeruginosa) Disulphides, Geosmin (flos-aquae), Disulphides, iso-propyl mercaptan(Wesenbergii)
The cells of microcystis are small – only a few microns in diameter – and lack individual mucilage sheaths. The cells are usually arranged in colonies that are initially spherical but become irregular or perforated over time. The cells may be grouped tightly or sparsely within the fine, colourless colonial mucilage. The colonies are free-floating and may be composed of clustered subcolonies. Smaller colonies are microscopic, while larger colonies may be viewed with the naked eye.
The cells are spherical or hemispherical after dividing. The protoplast is a pale blue-green, but overall the cells appear dark or brownish due to the presence of gas vesicles. This feature easily distinguishes microcystis from aphanocapsa under light microscopy.




Dolichospermum (formerly anabaena) is a cyanobacteria genus that is commonly found in freshwater phytoplankton assemblages. In nutrient-rich lakes it can form dense blooms. Superficially, Dolichospermum and anabaena look similar, but Dolichospermum can form gas vesicles, making it planktonic. Anabaena never forms gas vesicles and is usually associated with benthic environments.
Individual Dolichospermum cells are spherical, oval, or barrel-shaped
Under magnification Dolichospermum cells are dark brown and appear granular or mottled due to gas vesicles in the cells. Anabaena will have cells that are pale blue-green or blue-grey in colour.
The cells are joined together end to end to form long, unbranched filaments that are surrounded by clear, often transparent mucilage.
Depending on the species, the filaments can be straight, bent, coiled, or irregularly twisted, and may be solitary or aggregated into tangled clumps.
In addition to ordinary (vegetative) cells, the filaments may contain pale blue heterocytes and large, granular, thick-walled akinetes.
Heterocysts are specialised cells that convert dissolved nitrogen gas into ammonium that can be used for cell growth.
Akinetes are resting cells that are resistant to cold temperatures and other unfavourable environmental conditions, and can overwinter in lake sediments.
Dolichospermum blooms often form during warm, calm weather in lakes and ponds with relatively high nutrient concentrations (nitrogen or phosphorous) or low nitrogen to phosphorus ratios (N:P<15).
Recent work suggests that high total phosphorus (TP) or total Nitrogen (TN) concentrations are better predictors of bloom formation than N:P ratios. The gas vesicles in Dolichospermum cells provide a mechanism to move up and down in the water column, which increases access to nutrients and other growth factors. Because Dolichospermum is capable of converting dissolved nitrogen gas ammonium it can dominate blooms when inorganic nitrogen (ammonium, nitrate and nitrite) is limiting to other types of algae.
Nitrogen fixation requires a large amount of energy, so the relationship between nitrogen concentrations and Dolichospermum blooms is complicated; blooms can develop under both low and high inorganic nitrogen concentrations.
Dolichospermum blooms usually contain other types of Cyanobacteria, especially Aphanizomenon, Gloeotrichia, Microcystis and Woronichinia. Although Dolichospermum filaments are rarely consumed by zooplankton, the cells may be parasitized by viruses, bacteria or fungi. Tangled clumps of Dolichospermum may be colonised by stalked, filter feeding zooplankton.
Higher water temperatures and light appear to be associated with increased toxin production.
Earthy, musty
Metabolite produced – Geosmin
Dolichospermum (formerly anabaena) is a cyanobacteria genus that is commonly found in freshwater phytoplankton assemblages. In nutrient-rich lakes it can form dense blooms. Superficially, Dolichospermum and anabaena look similar, but Dolichospermum can form gas vesicles, making it planktonic. Anabaena never forms gas vesicles and is usually associated with benthic environments.
Individual Dolichospermum cells are spherical, oval, or barrel-shaped
Under magnification Dolichospermum cells are dark brown and appear granular or mottled due to gas vesicles in the cells. Anabaena will have cells that are pale blue-green or blue-grey in colour.
The cells are joined together end to end to form long, unbranched filaments that are surrounded by clear, often transparent mucilage.
Depending on the species, the filaments can be straight, bent, coiled, or irregularly twisted, and may be solitary or aggregated into tangled clumps.
In addition to ordinary (vegetative) cells, the filaments may contain pale blue heterocytes and large, granular, thick-walled akinetes.
Heterocysts are specialised cells that convert dissolved nitrogen gas into ammonium that can be used for cell growth.
Akinetes are resting cells that are resistant to cold temperatures and other unfavourable environmental conditions, and can overwinter in lake sediments.
Dolichospermum blooms often form during warm, calm weather in lakes and ponds with relatively high nutrient concentrations (nitrogen or phosphorous) or low nitrogen to phosphorus ratios (N:P<15).
Recent work suggests that high total phosphorus (TP) or total Nitrogen (TN) concentrations are better predictors of bloom formation than N:P ratios. The gas vesicles in Dolichospermum cells provide a mechanism to move up and down in the water column, which increases access to nutrients and other growth factors. Because Dolichospermum is capable of converting dissolved nitrogen gas ammonium it can dominate blooms when inorganic nitrogen (ammonium, nitrate and nitrite) is limiting to other types of algae.
Nitrogen fixation requires a large amount of energy, so the relationship between nitrogen concentrations and Dolichospermum blooms is complicated; blooms can develop under both low and high inorganic nitrogen concentrations.
Dolichospermum blooms usually contain other types of Cyanobacteria, especially Aphanizomenon, Gloeotrichia, Microcystis and Woronichinia. Although Dolichospermum filaments are rarely consumed by zooplankton, the cells may be parasitized by viruses, bacteria or fungi. Tangled clumps of Dolichospermum may be colonised by stalked, filter feeding zooplankton.
Higher water temperatures and light appear to be associated with increased toxin production.
Earthy, musty
Metabolite produced – Geosmin
**
**



**
**
**
**

Colonies more or less spherical or irregularly oval commonly composed of subcolonies, free-living, usually with a narrow, colourless enveloping mucilaginous layer.
In the centre of colonies is a system or radially and more or less parallel oriented, simple unbranched stalks, with split along their entire length after cell division; stalks thick, usually as wide as the width of the cells, commonly densely packed together and so causing radial lamellations of the central colonial mucilage; the stalk system sometimes indistinct in old subcolonies.
Cells rarely almost spherical, usually slightly elongate, widely oval, oval or obovoid, separating from one another immediately after division, but in old colonies very densely, radially aggregated in the peripheral layer.
**
**
**
**




**
**
**



**
**
**
**

**
**
**
**